Hybridization between wheat and rye was started in the 1870's. The English scientist A.S.Wilson made the first hybridization by crossing wheat as the female parent and rye as the male parent, and the hybrid was highly sterile. In 1888, the fertile seeds were contained by German breeder W. Rimpau from the sterile hybrid, due to the natural doubling of the F1 hybrid by low temperature.
Because of the good characters of triticale, geneticists and breeders have cultivated various wheat and rye hybrids through backcross and selfing during the past one hundred years. About 70% of the released wheat varieties around the world contain a pair of special chromosomes which we called the 1RS.1BL translocation chromosomes.
Wheat-rye 1RS.1BL translocations are centric translocations, with the short arm of wheat chromosome 1B being replaced by the short arm of rye chromosome 1R. Why this whole arm translocation could make such immense contributions to wheat production?
Researches at Dr. HAN Fangpu's lab from the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, revealed the centromere structure and function in the wheat-rye translocation lines.
They collected hundreds of 1RS.1BL translocation cultivars around the world to check the fine structure of the centromeres. Interestingly, they found that all 1RS.1BL translocations contained fused centromeres, half of the centromere was derived from wheat and the other half was derived from rye (Figure 1A).
Study on the rye specific centromere retrotransposon pAWCR.1 showed that the rye centromere retrotransposon was actively transcribed in 1RS.1BL translocations. The wheat CENH3 genes were located on the long arms of homologous group-1 chromosomes (Yuan et al, 2015, New Phytologist). CENH3 immunolocalization analysis demonstrated that the rye centromere DNA in the fusion centromere use the wheat CENH3 to perform function.
They also tried to answer the question that why only the fusion centromere is used in wheat breeding? They made efforts for many years to obtain 1RS.1BL translocations with the whole wheat centromere or the whole rye centromere to study its effect on gene expression and agronomic performance, but they screened thousands of materials and found that the wheat-rye translocations from homologous group-1 are all fusion centromere translocations.
With the invalidation of the resistance genes on 1RS as well as the increasing demands for wheat breeding, they introduced 200 rye varieties from the U.S. National Plant Germplasm System (NPGS) to make new wheat-rye hybridizations. Lots of amphidiploids, a complete set of wheat-rye disomic addition lines and telosomic addition lines have been obtained.
The new 1RS.1BL translocations are fusion centromere. In the progenies of the 1R monosomic addition line, the 1R chromosome could translocate to wheat homologous group-1 to group-7 chromosomes. Besides, many chromosome and centromere variation materials have been found, including centromere expansion and neocentromere formation (Guo et al, 2016, PLOS Genetics).
The mechanism of 1RS.1BL translocation can be analyzed by using anti-CENH3 ChIP-seq data and genomic information. The new translocations have been backcrossed, and several high stable generations are used for wheat breeding (Figure 1B).
The research was supported by the Ministry of Science and Technology of China, and the Chinese Academy of Sciences.
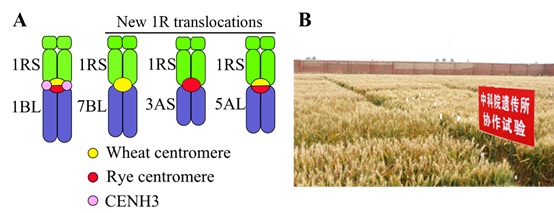
Figure 1. Centromere structure and function analysis and backcrossing in the 1RS.1BL translocations: (A) Schematics of the centromere structure and function in 1RS.1BL translocations and different centromere structures of 1R translocations. (B) Regional tests of the 1RS.1BL translocations (Image by IGDB)
















